- 2012年度
- 大脳皮質の進化の謎に迫る
(2013/03/29) - 活性型ビタミンDで小胞体ストレスを緩和しよう!
(2013/03/29) - Sweat Glands Grown from Newly Identified Stem Cells
(2013/03/29) - ~筋肉の恒常性と健康は相反するものなのか?~ 筋肉においてオートファジーが存在するが故にインスリン抵抗性が存在する
(2013/03/29) - たった1つの因子の抑制で様々な細胞が神経に!?
(2013/03/28) - 乳酸菌を取り込むと、細胞も若返る?! ~"人間、またしても発酵食品のお世話になる"の巻~
(2013/03/27) - 脳腫瘍における新しい遺伝子変異~エピジェネティク
(2013/03/27) - CCR2-dependentrecruitment of macrophages by tumor-educated mesenchymal stromal cells promotes tumor development and is mimicked by TNFα
(2013/03/25) - 補体C1qはWntシグナルの活性化を介して細胞を老化させる
(2013/03/19) - Speramtogonial Stem Cell Transplantation into Rhesus Testes Regenerates Spermatogenesis Producing Functional Sperm.
(2013/03/12) - 糖尿病薬剤による抗腫瘍効果
(2013/03/12) - 難病ALSの新たな原因遺伝子の発見
(2013/03/12) - 脊髄損傷にヒートショックプロテインが有効?
(2013/03/11) - 片頭痛患者では血管内皮前駆細胞が少ない?
(2013/03/11) - 筋幹細胞の静止状態はmiRNA-489により維持される
(2013/03/11) - ストレスに弱いってどういうこと?(心の病気にかかるメカニズムの一つ、「ストレス脆弱モデル」をネズミで再現)
(2013/03/09) - 核酸医薬は実現するか~筋強直性ジストロフィー治療の可能性~
(2013/03/09) - なくならないのは技がある!
(2013/03/08) - Down症候群のiPS細胞の染色体数を修正する
(2013/03/08) - 高品質なiPS細胞作製のキーファクターZscan4の同定
(2013/03/08) - Turning off the Neuron Death Pathway
(2013/03/07) - 新しい安全な分子標識-マルチ同位体画像質量分析法-が明らかにした幹細胞の不等分裂様式
(2013/03/07) - 重度脊髄損傷後に移植した神経幹細胞が非常に長く軸索を伸長し、シナプス結合した!
(2013/03/07) - 神経系前駆細胞を元気にして水頭症を治す!?
(2013/03/05) - FUS/TLSとTDP-43 二つのALS原因遺伝子の交差点
(2013/03/01) - 貪食細胞マクロファージが造血幹細胞を優しく包み込んで自己複製能の維持に貢献していた!?
(2013/02/27) - TALENs -新遺伝子改変技術が生命科学を変える!?-
(2013/02/27) - RESTタンパク質による遺伝子発現調節 ~遺伝子発現とシナプス機能~
(2013/02/25) - 腸に住んでいるある平凡な細菌によって大腸がんは引き起こされる!
(2013/02/25) - 癌抑制遺伝子p53の変異はメバロン酸経路を活性化することで、正常な乳腺の構造を失わせる
(2013/02/25) - 樹状細胞は制御性T細胞の恒常性をコントロールすることで多発性硬化症を寛解させる
(2013/02/25) - Schwann Cell Plasticity After Spinal Cord Injury Shown by Neural Crest Lineage Tracing
(2013/02/15) - エクソソームは、癌細胞の「飛び道具」!
(2013/02/08) - 老化したニッチでは筋肉幹細胞は静止状態を保てない
(2013/01/31) - 幹細胞を使った創薬開発
(2013/01/31) - 体細胞リプログラミングにおける遺伝子発現調節の解析からわかること-single cellで見てみようの巻-
(2013/01/31) - がん幹細胞発生のかぎを握るのは誰?-ユーイング肉腫がん幹細胞の解析を通じた検証-
(2013/01/31) - 小動物用PET(Positron Emission Tomography)で、ラットの脳梗塞巣を探知することができる [18F]BMS-PET
(2013/01/31) - 脳の神経ネットワークにおけるヤングパワー!
(2013/01/18) - アストロサイトの性格はどうやって決まる?
(2013/01/18) - 幹細胞の自己複製能を制御する因子とは?
(2013/01/18) - アルデヒドが真犯人!?DNA損傷と再生不良性貧血
(2012/12/18) - HIV-2の新しい定量法
(2012/12/18) - Japanese People's Preference for Place of End-of-Life Care and Death: A Population-Based Nationwide Survey
(2012/12/18) - がん細胞の死に際
(2012/12/18) - 癌幹細胞を眠りから目覚めさせる"Coco"
(2012/12/13) - 細胞接着分子のインテグリンが血液の幹細胞の維持を制御する
(2012/12/11) - 移植された神経幹細胞は免疫系にも作用する
(2012/12/11) - 血液がん克服にむけて!~JAK2阻害剤の薬剤耐性メカニズム解明~
(2012/12/04) - 癌進展を陰で操る支配者
(2012/11/30) - 幹細胞の2つの顔を暴け!!! 未分化性維持と特異的分化との狭間で...
(2012/11/30) - 脊髄損傷後の機能回復には自発的なリハビリが効果的
(2012/11/21) - もしあなたの歯が無くなってしまった時に...
(2012/11/15) - iPS細胞から血小板をつくる
(2012/10/30) - メラノーマのエキソソームで予後予測ができる?!
(2012/10/30) - Oligodendroglia Cells Can Do Much More Than an Insulator for Neuron
(2012/09/11) - APJは、心臓肥大のデュアル受容体として作用する
(2012/09/11) - 幹細胞医療;脳梗塞治療への挑戦
(2012/09/11) - 造血幹細胞の老化と若返り
(2012/09/11) - カロリー制限が筋肉を増やす? - トレーニング界の常識に挑戦する新たな"逆説"
(2012/09/11) - 癌幹細胞は治療標的にならない!?
(2012/08/24) - iPS細胞でC型肝炎ウイルス感染のモデルをつくる
(2012/08/09) - ES細胞、iPS細胞から内耳有毛細胞への分化誘導
(2012/08/09) - 造血幹細胞を冬眠させる細胞はなんと神経系の細胞だった!
(2012/07/06) - 個別化治療への障壁 ~多重人格なガンを克服せよ~
(2012/07/06) - 栄養のバランスが新しいニューロンを作り、体重や新陳代謝をコントロールする
(2012/05/11) - 脊髄不全損傷後におこる、残存神経ネットワークの代償機能
(2012/05/11) - "スーパーPTENマウス"
(2012/04/20) - 統合失調症iPS細胞研究が臨床研究になるために
(2012/04/20) - 発癌機序における"はじめの一歩"
(2012/04/06) - iPS細胞は脊髄損傷を治せるのか?
(2012/04/06)
- 大脳皮質の進化の謎に迫る
- 2011年度
- 2010年度
ホーム > 世界の幹細胞(関連)論文紹介 > 核酸医薬は実現するか~筋強直性ジストロフィー治療の可能性~
核酸医薬は実現するか~筋強直性ジストロフィー治療の可能性~
論文紹介著者

大塚 貴文(博士課程 1年)
GCOE RA
生理学教室
第一著者名・掲載雑誌・号・掲載年月
Thurman M. Wheeler/Nature. 488, 111-115, 02 August 2012
文献の英文表記:著者名・論文の表題・雑誌名・巻・号・ページ・発行年(西暦)
Wheeler TM, Leger AJ, Pandey SK, MacLeod AR, Nakamori M, Cheng SH, Wentworth BM, Bennett CF, Thornton CA. Targeting nuclear RNA for in vivo correction of myotonic dystrophy. Nature. 2012 Aug 2;488(7409):111-5.
論文解説
背景
核酸医薬とはDNAやRNAなどの核酸をその骨格にもつ医薬品で、アンチセンスやリボザイム、デコイ核酸など様々な手法が試みられており、いずれも特定の塩基配列やそれに結合するタンパク質を標的としているため、非常に特異性が高いという利点があります。
本論文で治療対象としている筋強直性ジストロフィー(DM1)は、筋細胞で発現するミオトニンプロテインキナーゼ(DMPK)遺伝子の3'非翻訳領域でCTG反復配列が増幅していることが原因とされています。CTG配列の増幅によって、転写産物であるmRNAが核内に蓄積し、MBNLやCUG-BPといったRNA結合タンパク質(※1)に結合し、クロライドチャネルや筋小胞体カルシウムATPアーゼといった筋肉の収縮・弛緩に重要なタンパク質遺伝子のRNAスプライシング(※2)の異常を引き起こすことが病態の原因であることが報告されています(図1)。
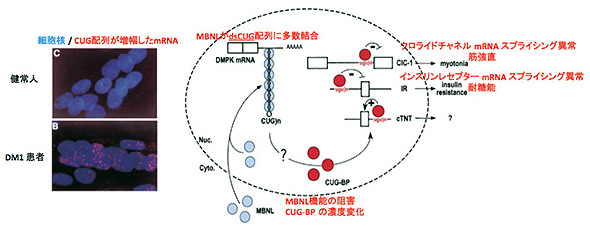
図1 DM1患者筋細胞核内での異常mRNAの蓄積とそれがもたらす病態
従来のような一つのタンパク質だけを標的とした医薬品ではこのような疾病の根本的治療は不可能でした。そこで、筆者らはアンチセンスオリゴヌクレオチド(※3)(ASO)と呼ばれる核酸医薬によって、核内に蓄積する有害なmRNAを分解してしまおうという戦略をとっています。ASOを用いたDM1の治療は、異常なmRNAのCUG反復配列部分に対するASOが既に試みられていますが、筋細胞への取り込みが非常に少ないため、生体に用いることができるレベルでは成功していませんでした。
方法
筆者らは、筋強直性ジストロフィー(DM1)の動物モデルとして、HSALRマウスを用いています。このマウスはhACTA遺伝子(筋細胞で恒常的に発現するヒト遺伝子)3'側CTG反復配列を220回に増幅したものをゲノム中に組込んだトランスジェニック動物(※4)で、筋細胞でCUG配列が増幅したmRNAが発現することによって、DM1同様の症状が現れます。
また、ASOは分解されやすく、骨格筋や心筋細胞には取り込まれにくいという難点がありましたが、筆者らはリボースのホスホジエステル結合をホスホロチオエート化し、さらにASO両端の5ヌクレオチドのリボース2位炭素をメトキシエチル化することでRNaseに対する安定性を向上させ、筋細胞への取り込みを促進しています(ただし、中心の10ヌクレオチドに関してはRNaseH1による標的mRNAの分解を誘導するため改変を加えていません。これをMOE gapmerと呼んでいます(図2))。
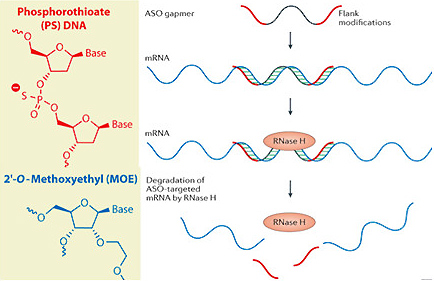
図2 ホスホロチオエート化とメトキシエチル化したASO(MOE gapmer)とその作用機序
これらの工夫によって皮下投与でも筋細胞にASOが届き、さらに投与後一年以上体内に残存することが示されています。
結果
筆者らはHSALRマウスにhACTAのmRNAに対するASO(MOE gapmer)を投与したところ、筋細胞の核内に蓄積した異常なmRNAが特異的に分解され、さらにクロライドチャネルや筋小胞体カルシウムATPアーゼのPre-mRNAスプライシングも正常化することを確認しました。また、筋電図測定によって強直性ジストロフィーに診られるミオトニー電位が解消することも明らかとなりました。さらに、これらの効果は、ASOを投与して1年後でも継続しており、安定性の向上も示されています(図3)。
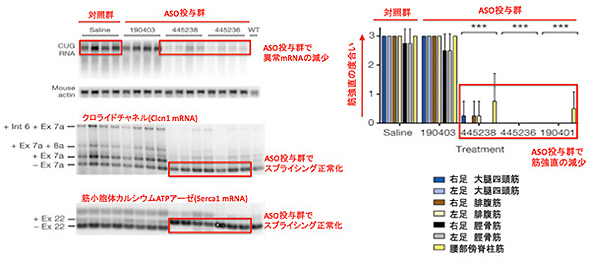
図3 ASOによる異常なmRNAの分解とスプライシング正常化、筋強直の解消
ただしASOによるmRNAの分解効果は、主に核内で働くRNaseH1という酵素に依存するため、細胞質などに標的mRNAがいる場合は分解効果が得られないことが示唆されています。
まとめ
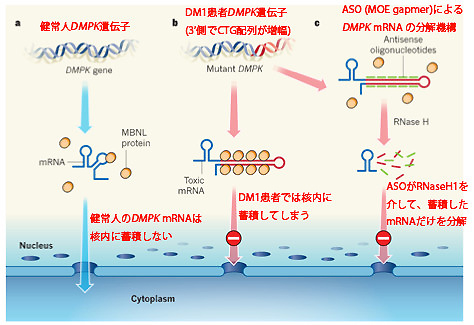
図4 ASOによるDM1治療のメカニズム
異常なmRNAが核内に蓄積することで引き起こされる筋強直性ジストロフィー(DM1)は、ASOによるRNaseH1を介したmRNAの分解に非常に感受性が高いことが示唆され、モデルマウスに対するASO皮下投与では高い治療効果が得られました(図4)。これはDM1同様に核内に異常なmRNAが蓄積することが原因となる疾患に対して応用できる可能性を示しています。
用語解説
- ※1 RNA結合タンパク質:
細胞内でmRNAと結合することでRNAスプライシングを制御したり、細胞質での分解を抑制するなど多様な機能をもつタンパク質の総称。細胞内での局在とターゲットmRNAへの結合位置によってその機能は変化する。 - ※2 RNAスプライシング:
核内でDNAがPre-mRNAへ転写された後、イントロンの除去とエクソンの再結合が行われる。選択的スプライシングでは、複数あるエクソンの一部はイントロンと共に選択的に除去される。 - ※3 アンチセンスオリゴヌクレオチド:
標的となるmRNAの一部と相補的な配列をもつ人工的に合成された核酸配列。 - ※4 トランスジェニック動物:
外部から人為的に特定の遺伝子を導入した遺伝子改変動物。
Copyright © Keio University. All rights reserved.