- 2012年度
- 2011年度
- 新たに判明 がんの転移を促進するメカニズム
(2012/03/23) - 神経発達と加齢における5-hmCを介したエピジェネティクス
(2012/03/23) - TACEを調節するiRhom2は、リステリア菌やLPSの反応により産生されるTNFを制御している
(2012/03/09) - HIV-2って何?
(2012/03/09) - 自殺遺伝子を持ったiPS細胞
(2012/03/09) - リハビリって神経幹細胞も殖やすんです!
(2012/02/24) - 癌幹細胞を制御するHippo pathway
(2012/02/10) - 吸血鬼が若い血を好むのには根拠があった?!~若い生き血でボケ防止~
(2012/02/10) - 癌幹細胞を特異的に標的とした治療法を開発できる可能性!?
(2012/01/27) - 骨の再生には、本来体を守る役割を持つはずのサイトカインは邪魔になる!?
(2012/01/27) - 骨格筋の老化は防げる?
(2012/01/13) - 意外に他力本願???他者の掘ったトンネルを行く癌細胞
(2012/01/13) - 新しいRNA間コミュニケーションのカタチ@筋肉
(2011/12/23) - 精子形成に必須なタンパク質Miwiによるトランスポゾンの発現抑制
(2011/12/23) - Dying well with dementia
(2011/12/09) - Recent insights into the epigenetic regulation of the hair follicle bulge stem cells
(2011/12/09) - ヒトiPS細胞から誘導した神経幹細胞における脳梗塞に対する移植治療の可能性
(2011/11/25) - 体細胞の再プログラム化を阻む"小さなRNA: miR-34"
(2011/11/25) - 薬剤性過敏症症候群 - DIHSがつなぐ薬疹とウイルスとの関連性
(2011/11/11) - 線維芽細胞より作製したドパミン作動性ニューロンは生体内において機能的であるのか?
(2011/11/11) - 終末分化した肝細胞から機能的な神経細胞への直接的な系統転換
(2011/10/28) - Nerves and T Cells Connect
(2011/10/28) - Rapid and robust generation of functional oligodendrocyte progenitor cells
(2011/10/28) - 脂肪細胞が発毛を促進する!?
(2011/10/14) - ADAM13はClass B Ephrinsの分解とWntシグナルの調節により頭部神経冠を誘導する
(2011/10/14) - 多能性の維持に働くchromatin remodeling複合体esBAF
(2011/09/30) - 造血幹細胞の維持にはp57が重要である
(2011/09/30) - IGF-II : 記憶力がよくなる分子!?
(2011/09/16) - 固形腫瘍に存在する間葉系幹細胞は癌幹細胞を増加させる
(2011/09/16) - 小腸は抑制性Th17細胞の宝庫
(2011/09/02) - 細胞周期を制御する新規noncoding RNA
(2011/09/02) - Sema3A play an important role in remyelination failure in multiple sclerosis
(2011/08/19) - Drosophila Sex lethal Gene initiates Female Development in Germline Progenitors
(2011/08/19) - Wnt signaling is a key pathway for regulation of Melanocyte stem cells.
(2011/08/05) - A step closer to understanding the heart
(2011/08/05) - 神経再生を阻む「死」のシグナル
(2011/07/25) - テロメラーゼの再活性化によりマウスの組織老化が回復する
(2011/07/25) - 新遺伝子「Glis1」により、安全なiPS細胞を高効率に作製可能
(2011/07/08) - 幹細胞の"状態"をつくりだす細胞外環境
(2011/07/08) - 毛包幹細胞、色素幹細胞を維持
(2011/06/24) - BCL6を標的とした白血病の新たな治療戦略
(2011/06/24) - 自家移植におけるiPS細胞の免疫応答について
(2011/06/03) - ヒト疾患iPS細胞のウィルソン病への応用
(2011/06/03) - FOP(進行性骨化性線維異形成症)の異所性骨化部の起源は?
(2011/04/22) - 非対称分裂がNotchシグナルの活性化を介して皮膚の分化を促進する
(2011/04/22) - ショウジョウバエの腸管幹細胞の増殖は活性酸素により制御される
(2011/04/22) - 線維芽細胞からの直接的なエピブラストステムセルの誘導
(2011/04/08) - 抗リウマチ薬DHODH阻害剤はメラノーマの進展を抑える
(2011/04/08) - 癌再発の指標になる幹細胞
(2011/04/08)
- 新たに判明 がんの転移を促進するメカニズム
- 2010年度
ホーム > 世界の幹細胞(関連)論文紹介 > HIV-2って何?
HIV-2って何?
論文紹介著者

山崎 さやか(博士課程 1年)
GCOE RA
微生物学・免疫学教室
第一著者名・掲載雑誌・号・掲載年月
Thushan I. de Silva/Trends in Microbiology Vol.16 No.12 588-595 2008年10月
文献の英文表記:著者名・論文の表題・雑誌名・巻・号・ページ・発行年(西暦)
Thushan I. de Silva1, Matthew Cotten1 and Sarah L. Rowland-Jones. HIV-2: the forgotten AIDS virus. Trends in Microbiology Vol.16 No.12.588-595,2008
論文解説
はじめに
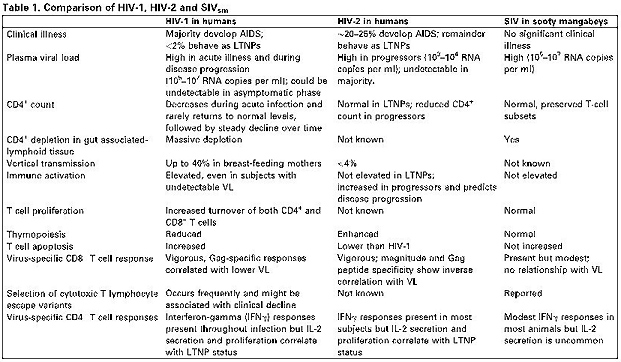
HIV-2はHIV-1の発見より数年後に発見されたウイルスであり、AIDSをごく少人数にしか発症しないウイルスである。なぜHIV-2が多くの感染者に対して症状を発症しないのか、これについてはまだよくわかっていない。しかし、最近の研究ではHIV-2に感染すると宿主の免疫反応が強くなるから、あるいはウイルスの複製能力がもともと弱いからAIDSが発症しないのではないかということが提唱されている。
HIV-2の研究をすることはどんな意味があるのか?
HIV-1ワクチン開発は未だに成功していない。大きな原因とは、感染あるいは病気の進行から守ろうとする宿主の免疫応答とウイルス機構についてよくわかっていないからである。これをよく知るためには、サル免疫不全ウイルスを調べること、またHIV-1感染者のうち長期に渡って病状が進行しない人たち(LTNPs)を調べることが一番の方法である。しかし、これらの研究は検体数が少なく、なかなか進めることができない。そこで、HIV-1によく似たウイルスであるHIV-2の研究が必要となってくるのである。HIV-2感染者の中にはLTNPsがたくさんおり、HIV-2の研究をすることによってHIVの免疫学的病体学を理解する価値ある洞察を得ることができるのである。
HIV-2起源とその分布
HIV-2は西アフリカのテナガザルから見つかったSIVとよく似ているので、これが起源だと考えられている。HIV-2感染者のうち多くはHIV-2西アフリカにいる。多くの国では、HIV-1の感染者が増加している一方で、HIV-2感染者は減少している。
HIV-2に感染したらどうなるのか?
HIVに感染した人のうち5年後にAIDSを発症していない人は、HIV-1が67%であるのに対しHIV-2は100%である。つまりCD4+T細胞の減りがHIV-1感染者に比べてかなり遅い。HIV-2の感染者のうち死亡するリスクがあるのは、感染していない人より単に2倍高いだけで、HIV-1感染者の10倍から20倍低いという報告もある。臨床的な特徴としてはHIV-2の感染者はカポジ肉腫をほとんどの人から観察されないことをのぞいてはHIV-1と同じである。
HIV-2は感染力が弱いウイルスなの?
HIV-2の感染者がほとんどAIDSにならないことから、ウイルスの複製能力が低い、宿主の免疫応答が高まる、あるいは両方ではないかと考えられている。症状が出ていない段階でのウイルス量(VL)と血漿中のRNA量は、HIV-2感染者はHIV-1感染者の30倍ほど低い。同じHIV-1とHIV-2感染者のうちCD4+T細胞量が同じぐらいの人を比べてみると、プロウイルスの量は同じであるが、HIV-1感染者の方が血漿中のVLは高い。これらのことから、HIV-2のプロウイルスはより強い規制を受けているのではないかと考えられる。実際、HIV-2は転写領域よりもヘテロクロマチン組み込まれやすいという報告もされている。
HIV-2に感染した時の宿主の免疫応答
ヒトではない霊長類の研究ではHIV-1の感染から守ろうとしてTRIM5αの経路が発現するがわかっている。臨床的コホート研究で、様々なTRIM5αは感染HIVの感染するリスクをさげているがわかった。シチジン脱アミノ酵素であるAPOBECファミリーもまたウイルスを制限するのだが、HIV-2もまた感受性がAPOBEC3Gに感受性がある。
HIV-1とHIV-2の二重感染
HIV-2の感染者はHIV-1の感染をしにくいという報告がある。しかし、西アフリカの報告ではHIV-2の感染者のほうがHIV-1の感染のリスクを高めているという報告もされている。これはまだ生物学的なものなのか、あるいは行動学的なのかはわかっておらず、またこれを証明するのは難しい。In vitro の研究ではHIV-2の感染はHIV-1の感染を防いでくれているという結果が出ているが、だからといって完全に感染を防いでくれるわけではないので「自然のワクチン」というわけにはいかない。
HIV-2の治療
HIV-2に感染者のなかで抗レトロウイルスの治療を受けている人はすくない。また、HIV-2の感染の治療のガイドラインをきちんとできていない。HIV-1の治療と同様なものが求められている。
HIV-2研究の今後は?
HIV-1の感染とは異なり、免疫応答で出てきたNab、これはHIV-1のワクチン開発のためによく研究されているものなのだが、これはHIV-2でもよく見られる。また、HIV-2の免疫応答によってもHIV-1の感染のリスクがへることは、HIVのワクチンの開発の鍵になるかもしれない。もちろん、HIV-2のワクチンの開発もかなり意味のあることである。約100万人ものHIV-2の感染者は西アフリカにいてARTの治療を受けることはできていないからだ。予防と治療目的のHIV-2ワクチンは、HIV-2の感染のリスクがある地域に役立つのみならずHIV-1の開発の役に立つだろう。
用語解説
- ※1 LTNPs
long-term non- progressors - ※2 VL
viral load - ※3 コホート
分析疫学における手法の1つであり、特定の要因に曝露した集団と曝露していない集団を一定期間追跡し、研究対象となる疾病の発生率を比較することで、要因と疾病発生の関連を調べる観察的研究である。
Copyright © Keio University. All rights reserved.