- 2012年度
- 2011年度
- 新たに判明 がんの転移を促進するメカニズム
(2012/03/23) - 神経発達と加齢における5-hmCを介したエピジェネティクス
(2012/03/23) - TACEを調節するiRhom2は、リステリア菌やLPSの反応により産生されるTNFを制御している
(2012/03/09) - HIV-2って何?
(2012/03/09) - 自殺遺伝子を持ったiPS細胞
(2012/03/09) - リハビリって神経幹細胞も殖やすんです!
(2012/02/24) - 癌幹細胞を制御するHippo pathway
(2012/02/10) - 吸血鬼が若い血を好むのには根拠があった?!~若い生き血でボケ防止~
(2012/02/10) - 癌幹細胞を特異的に標的とした治療法を開発できる可能性!?
(2012/01/27) - 骨の再生には、本来体を守る役割を持つはずのサイトカインは邪魔になる!?
(2012/01/27) - 骨格筋の老化は防げる?
(2012/01/13) - 意外に他力本願???他者の掘ったトンネルを行く癌細胞
(2012/01/13) - 新しいRNA間コミュニケーションのカタチ@筋肉
(2011/12/23) - 精子形成に必須なタンパク質Miwiによるトランスポゾンの発現抑制
(2011/12/23) - Dying well with dementia
(2011/12/09) - Recent insights into the epigenetic regulation of the hair follicle bulge stem cells
(2011/12/09) - ヒトiPS細胞から誘導した神経幹細胞における脳梗塞に対する移植治療の可能性
(2011/11/25) - 体細胞の再プログラム化を阻む"小さなRNA: miR-34"
(2011/11/25) - 薬剤性過敏症症候群 - DIHSがつなぐ薬疹とウイルスとの関連性
(2011/11/11) - 線維芽細胞より作製したドパミン作動性ニューロンは生体内において機能的であるのか?
(2011/11/11) - 終末分化した肝細胞から機能的な神経細胞への直接的な系統転換
(2011/10/28) - Nerves and T Cells Connect
(2011/10/28) - Rapid and robust generation of functional oligodendrocyte progenitor cells
(2011/10/28) - 脂肪細胞が発毛を促進する!?
(2011/10/14) - ADAM13はClass B Ephrinsの分解とWntシグナルの調節により頭部神経冠を誘導する
(2011/10/14) - 多能性の維持に働くchromatin remodeling複合体esBAF
(2011/09/30) - 造血幹細胞の維持にはp57が重要である
(2011/09/30) - IGF-II : 記憶力がよくなる分子!?
(2011/09/16) - 固形腫瘍に存在する間葉系幹細胞は癌幹細胞を増加させる
(2011/09/16) - 小腸は抑制性Th17細胞の宝庫
(2011/09/02) - 細胞周期を制御する新規noncoding RNA
(2011/09/02) - Sema3A play an important role in remyelination failure in multiple sclerosis
(2011/08/19) - Drosophila Sex lethal Gene initiates Female Development in Germline Progenitors
(2011/08/19) - Wnt signaling is a key pathway for regulation of Melanocyte stem cells.
(2011/08/05) - A step closer to understanding the heart
(2011/08/05) - 神経再生を阻む「死」のシグナル
(2011/07/25) - テロメラーゼの再活性化によりマウスの組織老化が回復する
(2011/07/25) - 新遺伝子「Glis1」により、安全なiPS細胞を高効率に作製可能
(2011/07/08) - 幹細胞の"状態"をつくりだす細胞外環境
(2011/07/08) - 毛包幹細胞、色素幹細胞を維持
(2011/06/24) - BCL6を標的とした白血病の新たな治療戦略
(2011/06/24) - 自家移植におけるiPS細胞の免疫応答について
(2011/06/03) - ヒト疾患iPS細胞のウィルソン病への応用
(2011/06/03) - FOP(進行性骨化性線維異形成症)の異所性骨化部の起源は?
(2011/04/22) - 非対称分裂がNotchシグナルの活性化を介して皮膚の分化を促進する
(2011/04/22) - ショウジョウバエの腸管幹細胞の増殖は活性酸素により制御される
(2011/04/22) - 線維芽細胞からの直接的なエピブラストステムセルの誘導
(2011/04/08) - 抗リウマチ薬DHODH阻害剤はメラノーマの進展を抑える
(2011/04/08) - 癌再発の指標になる幹細胞
(2011/04/08)
- 新たに判明 がんの転移を促進するメカニズム
- 2010年度
ホーム > 世界の幹細胞(関連)論文紹介 > 多能性の維持に働くchromatin remodeling複合体esB...
多能性の維持に働くchromatin remodeling複合体esBAF
論文紹介著者

木下 泰輔(博士課程 1年)
GCOE RA
発生・分化生物学教室
第一著者名・掲載雑誌・号・掲載年月
DLena Ho/Nature cell biology, 2011 Jul 24;13(8): 903-913
文献の英文表記:著者名・論文の表題・雑誌名・巻・号・ページ・発行年(西暦)
Lena Ho, Erik L. Miller, Jehnna L. Ronan, Wen Qi Ho, Raja Jothi and Gerald R. Crabtree. esBAF facilitates pluripotency by conditioning the genome for LIF/STAT3 signalling and by regulating polycomb function. Nature Cell Biology. 13(8):903-913, 2011
論文解説
マウスES細胞やiPS細胞といった多能性幹細胞の未分化性を維持するためには、LIF(Leukemia Inhibitory Factor、白血病阻害因子)というサイトカインが必要であることが知られています。LIFは幾つかのシグナル経路を介して、多能性維持に重要な転写因子の発現を誘導していることが明らかになっています。一方で、転写活性化においては、転写活性化因子がDNAへ結合するためにクロマチン構造(※1)の状態の調節も重要となってきます。
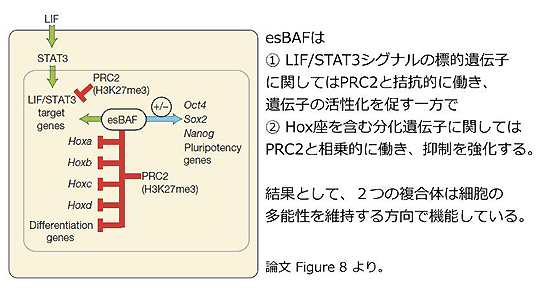
今回紹介する論文では、ES細胞で見られるクロマチン構造を調節する複合体esBAFとそのサブユニットBrg1が、LIFシグナルの下流の転写因子STAT3が多能性に関連する遺伝子に結合し易いようにゲノムを整えること、またPolycomb(PcG)複合体(※2)とあるときは拮抗的に・あるときは相乗的に働くことで、多能性の維持に寄与していることを報告しています。
筆者らは薬剤(4-hydroxytamoxifen、4-OHT)投与によりBrg1を任意に欠損させることのできるES細胞を用いて、Brg1およびesBAFによる直接的な作用を評価しています。
まず、ES細胞においてBrg1とSTAT3のゲノムへの結合箇所が広範に重複していることから、これらが転写的にも同様の遺伝子集団を制御しているかどうかの検討を行いました。マイクロアレイデータ解析により、両者はかなりの数の共通した遺伝子を抑制したり活性化させたりしていることが示唆されました。この活性化される遺伝子群の中には、多能性に必須な遺伝子もいくつか含まれていました。
次にES細胞をLIFが欠乏した状態からLIF存在下に戻すという実験を行ったところ、野生型ES細胞で見られたLIFに応答した遺伝子の再活性化が、Brg1欠損ES細胞では一般的に損なわれていました。さらにChIP解析により、Brg1欠損細胞では遺伝子へのSTAT3の結合が障害されていることが明らかになりました。
一方で、Brg1はLIFシグナリングとは関係ない遺伝子も数多く制御しています。観察されたSTAT3の遺伝子への動員におけるBrg1の働きを評価するため、STAT3の増強によりBrg1欠損細胞でおきる自己複製の異常をレスキューできるか調べました。結果、STAT3の強制発現はBrg1欠損による自己複製異常を部分的に緩和しました。
さらに、遺伝子への結合の容易さをDNaseI高感度アッセイで評価したところ、Brg1の制御を受けるSTAT3部位は制御を受けないSTAT3部位に比べてその容易さが低いという結果が得られました。Brg1を欠損させると、制御を受けるSTAT3部位はさらに容易さを低下させることから、Brg1はSTAT3が標的部位に結合しやすくなるようにクロマチン構造の調節を行うことが示唆されました。
ここでBrg1欠損細胞におけるヒストン修飾を調べたところ、Brg1に制御されるSTAT3標的遺伝子のプロモーター領域において、活性型のしるしであるH3K4me3やH4Acに変化は見られなかったのに対し、抑制的な調節のしるしであるH3K27me3の増加が観察されました。
H3K27me3はpolycomb repressive complex2(PRC2)という複合体により付与されることから、筆者らは最後にesBAFとPcG複合体の機能的な相互作用について検討しました。Hox遺伝子座において、Brg1欠損ES細胞でH3K27me3の顕著な減少が見られたことから、Brg1はPcGによる抑制化を手助けするように機能し、ES細胞が分化するのを防いでいることが示唆されました。一方、STAT3の標的部位について再度詳しく解析し、Brg1欠損細胞では全体的にH3K27me3の著しい上昇が見られること、PRC2の構成タンパクSuz12をshRNAで減少させることで上昇したH3K27me3のレベルが野生型のレベルまで戻ることを確認しました。
DNA配列の変化を伴わず、クロマチンの状態変化によって調節される遺伝子の発現制御はエピジェネティクスと呼ばれていますが、幹細胞研究においても、このエピジェネティクスの視点から多能性を捉えるということに関して注目が集められ、研究が進められています。今回紹介した論文もそうした流れのなかに位置しており、幹細胞の多能性理解のために、いろいろな側面からのさらなる研究の進展が期待されます。
用語解説
- ※1 クロマチン構造
細胞の核内において、DNAがヒストンタンパク質に巻き付いて形成している構造。
ヒストンにはH1、H2A、H2B、H3、H4の5種類があり、これらヒストンが化学的修飾(メチル化やアセチル化など)を受けることでクロマチン構造が変化する。 - ※2 Polycomb(PcG)複合体
ヒストンを化学的に修飾するタンパク質複合体で、遺伝子の発現を抑制化方向に制御する。
遺伝子の発現を活性化方向に制御するタンパク質複合体としては、Trithorax(TrxG)複合体がある。
Copyright © Keio University. All rights reserved.