- 2012年度
- 2011年度
- 新たに判明 がんの転移を促進するメカニズム
(2012/03/23) - 神経発達と加齢における5-hmCを介したエピジェネティクス
(2012/03/23) - TACEを調節するiRhom2は、リステリア菌やLPSの反応により産生されるTNFを制御している
(2012/03/09) - HIV-2って何?
(2012/03/09) - 自殺遺伝子を持ったiPS細胞
(2012/03/09) - リハビリって神経幹細胞も殖やすんです!
(2012/02/24) - 癌幹細胞を制御するHippo pathway
(2012/02/10) - 吸血鬼が若い血を好むのには根拠があった?!~若い生き血でボケ防止~
(2012/02/10) - 癌幹細胞を特異的に標的とした治療法を開発できる可能性!?
(2012/01/27) - 骨の再生には、本来体を守る役割を持つはずのサイトカインは邪魔になる!?
(2012/01/27) - 骨格筋の老化は防げる?
(2012/01/13) - 意外に他力本願???他者の掘ったトンネルを行く癌細胞
(2012/01/13) - 新しいRNA間コミュニケーションのカタチ@筋肉
(2011/12/23) - 精子形成に必須なタンパク質Miwiによるトランスポゾンの発現抑制
(2011/12/23) - Dying well with dementia
(2011/12/09) - Recent insights into the epigenetic regulation of the hair follicle bulge stem cells
(2011/12/09) - ヒトiPS細胞から誘導した神経幹細胞における脳梗塞に対する移植治療の可能性
(2011/11/25) - 体細胞の再プログラム化を阻む"小さなRNA: miR-34"
(2011/11/25) - 薬剤性過敏症症候群 - DIHSがつなぐ薬疹とウイルスとの関連性
(2011/11/11) - 線維芽細胞より作製したドパミン作動性ニューロンは生体内において機能的であるのか?
(2011/11/11) - 終末分化した肝細胞から機能的な神経細胞への直接的な系統転換
(2011/10/28) - Nerves and T Cells Connect
(2011/10/28) - Rapid and robust generation of functional oligodendrocyte progenitor cells
(2011/10/28) - 脂肪細胞が発毛を促進する!?
(2011/10/14) - ADAM13はClass B Ephrinsの分解とWntシグナルの調節により頭部神経冠を誘導する
(2011/10/14) - 多能性の維持に働くchromatin remodeling複合体esBAF
(2011/09/30) - 造血幹細胞の維持にはp57が重要である
(2011/09/30) - IGF-II : 記憶力がよくなる分子!?
(2011/09/16) - 固形腫瘍に存在する間葉系幹細胞は癌幹細胞を増加させる
(2011/09/16) - 小腸は抑制性Th17細胞の宝庫
(2011/09/02) - 細胞周期を制御する新規noncoding RNA
(2011/09/02) - Sema3A play an important role in remyelination failure in multiple sclerosis
(2011/08/19) - Drosophila Sex lethal Gene initiates Female Development in Germline Progenitors
(2011/08/19) - Wnt signaling is a key pathway for regulation of Melanocyte stem cells.
(2011/08/05) - A step closer to understanding the heart
(2011/08/05) - 神経再生を阻む「死」のシグナル
(2011/07/25) - テロメラーゼの再活性化によりマウスの組織老化が回復する
(2011/07/25) - 新遺伝子「Glis1」により、安全なiPS細胞を高効率に作製可能
(2011/07/08) - 幹細胞の"状態"をつくりだす細胞外環境
(2011/07/08) - 毛包幹細胞、色素幹細胞を維持
(2011/06/24) - BCL6を標的とした白血病の新たな治療戦略
(2011/06/24) - 自家移植におけるiPS細胞の免疫応答について
(2011/06/03) - ヒト疾患iPS細胞のウィルソン病への応用
(2011/06/03) - FOP(進行性骨化性線維異形成症)の異所性骨化部の起源は?
(2011/04/22) - 非対称分裂がNotchシグナルの活性化を介して皮膚の分化を促進する
(2011/04/22) - ショウジョウバエの腸管幹細胞の増殖は活性酸素により制御される
(2011/04/22) - 線維芽細胞からの直接的なエピブラストステムセルの誘導
(2011/04/08) - 抗リウマチ薬DHODH阻害剤はメラノーマの進展を抑える
(2011/04/08) - 癌再発の指標になる幹細胞
(2011/04/08)
- 新たに判明 がんの転移を促進するメカニズム
- 2010年度
ホーム > 世界の幹細胞(関連)論文紹介 > 毛包幹細胞、色素幹細胞を維持
毛包幹細胞、色素幹細胞を維持
論文紹介著者

外山 弘文(博士課程 2年)
GCOE RA
発生・分化生物学教室
第一著者名・掲載雑誌・号・掲載年月
Shintaro Tanimura/Cell Stem Cell. 2011 Feb 4;8(2):177-87.
文献の英文表記:著者名・論文の表題・雑誌名・巻・号・ページ・発行年(西暦)
Tanimura S, Tadokoro Y, Inomata K, Binh NT, Nishie W, Yamazaki S, Nakauchi H, Tanaka Y, McMillan JR, Sawamura D, Yancey K, Shimizu H, Nishimura EK. Hair follicle stem cells provide a functional niche for melanocyte stem cells. Cell Stem Cell. 2011 Feb 4;8(2):177-87.
論文解説
1.毛の中の幹細胞―毛包幹細胞と色素幹細胞
今回は毛の幹細胞についての論文を説明します。幹細胞は体中の様々な組織において存在が報告されていますが、毛にも幹細胞が存在することがわかってきました。実は、私たちの毛は体の外に見えている部分だけでなく、皮膚の表面の奥深くから生えています。この皮膚で隠れていて普段見えない部分のバルジと呼ばれる領域に毛の幹細胞である毛包幹細胞(※1)と色素幹細胞(※2)は存在します(図1)。毛包幹細胞は毛を形作る細胞である角化細胞の元の細胞として機能し、色素幹細胞は毛の色をなす色素細胞を生み出します。

2.毛包幹細胞に存在する17型コラーゲンの重要性
筆者らはバルジの幹細胞において17型コラーゲンが重要な役割を示すと考え、この分子に着目しました。17型コラーゲンは基底膜(※3)に貫通する形で存在し、基底膜の上に並ぶ基底細胞を基底膜に固定する役割があります。筆者らは野生型マウスの毛を調べることで、毛包幹細胞に17型コラーゲンが存在することを示しました。また、Col17a1遺伝子を欠損したノックアウトマウスを作製し、17型コラーゲンの機能を探索しました。このマウスは17型コラーゲンの構成要素であるコラーゲン17A1を持っていないため、17型コラーゲンが機能しないようになっています。このCol17a1ノックアウトマウスは一見すると通常の野生型マウスと同じように生まれてきました。しかし時を追って観察していると、体の毛がだんだん脱色していき、ついには全身の毛が抜け落ちてしまいました(図2)。この原因を調べると、興味深いことにCol17a1ノックアウトマウスの毛において、毛の脱色が起きる前の生後12週辺りに色素幹細胞が徐々に異常な形態に変わっていき、最終的には欠如してしまうことがわかりました。
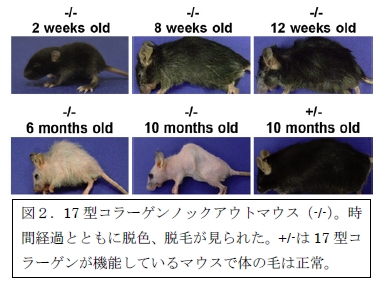
一方、毛包幹細胞についても調べると、野生型マウスと比べてCol17a1ノックアウトマウスでは生後8週以降に毛包幹細胞の数が顕著に減少していることがわかりました。一般的に、幹細胞は静止状態と未分化性を維持することが重要で、細胞分裂が止まった状態にあって細胞の形態や機能を変えていく細胞系列の大元に当たります。また、いわゆる自己複製能によって細胞分裂により自分自身のコピーを作ると同時に、他の細胞を形作っていきます(具体的には、毛包幹細胞の場合は毛の角質細胞を供給します)。しかし、Col17a1ノックアウトマウスの毛包幹細胞はこれらの幹細胞としての重要な性質を失っていました。すなわち、毛包幹細胞が幹細胞の性質を失うことで枯渇していき、徐々に毛を形作ることができなくなり、最終的に脱毛が生じたと考えられました。
これらの結果を考察すると以下のようにまとめられます。
(1) Col17a1ノックアウトマウスでは生後8週頃より毛包幹細胞が幹細胞としての性質を失い、成長とともに枯渇する。また、色素幹細胞は生後12週頃から異常な形態を示し始め、徐々に枯渇する。これらの原因により、毛の脱色と脱毛が見られる。
(2) 毛包幹細胞に存在する17型コラーゲンが毛包幹細胞の維持に重要であり、色素幹細胞の維持にも関与している。
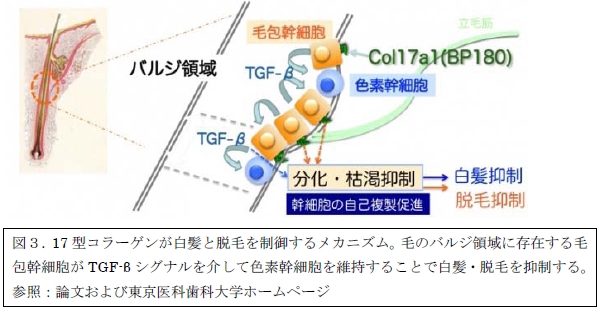
さらに、筆者は毛包幹細胞と色素幹細胞の関係において、毛包幹細胞が生じるTGF-βシグナル(※4)という伝達経路が重要であることを発見し、以下のモデルを提唱していました。つまり、毛包幹細胞は17型コラーゲンによって毛のバルジ領域に維持され、TGF-βを出すことによって色素幹細胞を維持していると考えられます。これまで様々な幹細胞において、ニッチと呼ばれる幹細胞周囲の微小な環境が幹細胞の維持に重要であることがわかっていますが、毛の幹細胞の制御の仕組みについては不明でした。今回の研究によって、毛包幹細胞がTGF-βシグナルを介して色素幹細胞のニッチ細胞として機能することが新たに提唱されました。幹細胞のニッチの制御機構を考えるうえで非常に興味深い発見です。今後は、白髪・脱毛のメカニズムのさらなる解明や将来の新たな治療の可能性が期待されます。
用語解説
- ※1 毛包幹細胞
毛を形作る角化細胞を供給する源の細胞。 - ※2 色素幹細胞
毛の色を呈する色素細胞を供給する源の細胞。 - ※3 基底膜
上皮細胞(皮膚では体の表面を覆う表皮)とその深部の間質細胞(皮膚など体のあらゆる構造の支持をする細胞)の間に存在する膜。膜上部に接して並ぶ基底細胞(皮膚の場合は表皮基底細胞)を支持する。 - ※4 TGF-βシグナル
細胞の増殖や分化(幹細胞を始めとした未熟な細胞が形態を変えて成熟すること)を制御するタンパク質による伝達経路。免疫反応やがんなど様々な病気において機能する。
Copyright © Keio University. All rights reserved.