- 2012年度
- 2011年度
- 新たに判明 がんの転移を促進するメカニズム
(2012/03/23) - 神経発達と加齢における5-hmCを介したエピジェネティクス
(2012/03/23) - TACEを調節するiRhom2は、リステリア菌やLPSの反応により産生されるTNFを制御している
(2012/03/09) - HIV-2って何?
(2012/03/09) - 自殺遺伝子を持ったiPS細胞
(2012/03/09) - リハビリって神経幹細胞も殖やすんです!
(2012/02/24) - 癌幹細胞を制御するHippo pathway
(2012/02/10) - 吸血鬼が若い血を好むのには根拠があった?!~若い生き血でボケ防止~
(2012/02/10) - 癌幹細胞を特異的に標的とした治療法を開発できる可能性!?
(2012/01/27) - 骨の再生には、本来体を守る役割を持つはずのサイトカインは邪魔になる!?
(2012/01/27) - 骨格筋の老化は防げる?
(2012/01/13) - 意外に他力本願???他者の掘ったトンネルを行く癌細胞
(2012/01/13) - 新しいRNA間コミュニケーションのカタチ@筋肉
(2011/12/23) - 精子形成に必須なタンパク質Miwiによるトランスポゾンの発現抑制
(2011/12/23) - Dying well with dementia
(2011/12/09) - Recent insights into the epigenetic regulation of the hair follicle bulge stem cells
(2011/12/09) - ヒトiPS細胞から誘導した神経幹細胞における脳梗塞に対する移植治療の可能性
(2011/11/25) - 体細胞の再プログラム化を阻む"小さなRNA: miR-34"
(2011/11/25) - 薬剤性過敏症症候群 - DIHSがつなぐ薬疹とウイルスとの関連性
(2011/11/11) - 線維芽細胞より作製したドパミン作動性ニューロンは生体内において機能的であるのか?
(2011/11/11) - 終末分化した肝細胞から機能的な神経細胞への直接的な系統転換
(2011/10/28) - Nerves and T Cells Connect
(2011/10/28) - Rapid and robust generation of functional oligodendrocyte progenitor cells
(2011/10/28) - 脂肪細胞が発毛を促進する!?
(2011/10/14) - ADAM13はClass B Ephrinsの分解とWntシグナルの調節により頭部神経冠を誘導する
(2011/10/14) - 多能性の維持に働くchromatin remodeling複合体esBAF
(2011/09/30) - 造血幹細胞の維持にはp57が重要である
(2011/09/30) - IGF-II : 記憶力がよくなる分子!?
(2011/09/16) - 固形腫瘍に存在する間葉系幹細胞は癌幹細胞を増加させる
(2011/09/16) - 小腸は抑制性Th17細胞の宝庫
(2011/09/02) - 細胞周期を制御する新規noncoding RNA
(2011/09/02) - Sema3A play an important role in remyelination failure in multiple sclerosis
(2011/08/19) - Drosophila Sex lethal Gene initiates Female Development in Germline Progenitors
(2011/08/19) - Wnt signaling is a key pathway for regulation of Melanocyte stem cells.
(2011/08/05) - A step closer to understanding the heart
(2011/08/05) - 神経再生を阻む「死」のシグナル
(2011/07/25) - テロメラーゼの再活性化によりマウスの組織老化が回復する
(2011/07/25) - 新遺伝子「Glis1」により、安全なiPS細胞を高効率に作製可能
(2011/07/08) - 幹細胞の"状態"をつくりだす細胞外環境
(2011/07/08) - 毛包幹細胞、色素幹細胞を維持
(2011/06/24) - BCL6を標的とした白血病の新たな治療戦略
(2011/06/24) - 自家移植におけるiPS細胞の免疫応答について
(2011/06/03) - ヒト疾患iPS細胞のウィルソン病への応用
(2011/06/03) - FOP(進行性骨化性線維異形成症)の異所性骨化部の起源は?
(2011/04/22) - 非対称分裂がNotchシグナルの活性化を介して皮膚の分化を促進する
(2011/04/22) - ショウジョウバエの腸管幹細胞の増殖は活性酸素により制御される
(2011/04/22) - 線維芽細胞からの直接的なエピブラストステムセルの誘導
(2011/04/08) - 抗リウマチ薬DHODH阻害剤はメラノーマの進展を抑える
(2011/04/08) - 癌再発の指標になる幹細胞
(2011/04/08)
- 新たに判明 がんの転移を促進するメカニズム
- 2010年度
ホーム > 世界の幹細胞(関連)論文紹介 > 脂肪細胞が発毛を促進する!?
脂肪細胞が発毛を促進する!?
論文紹介著者

吉田 和恵(博士課程 3年)
GCOE RA
皮膚科学教室
第一著者名・掲載雑誌・号・掲載年月
Eric Festa/Cell. 146(5):761-71. 2011 Sep
文献の英文表記:著者名・論文の表題・雑誌名・巻・号・ページ・発行年(西暦)
Eric Festa, Jackie Fretz, Ryan Berry, Barbara Schmidt, Matthew Rodeheffer, Mark Horowitz, and Valerie Horsley. Adipocyte Lineage Cells Contribute to the Skin Stem Cell Niche to Drive Hair Cycling. Cell. 146(5):761-71. 2011
論文解説
毛には、抜けては生えることを繰り返す生え変わりのサイクル(毛周期)があります。毛周期は,成長期(anagen)、退行期(catagen)、休止期(telogen)の順を繰り返します。ヒトの髪の毛の場合では、毛は数年間細胞分裂を繰り返して成長を続け、その後退行期に移行すると細胞分裂は止まり、2~3週間かけて退化を起こし、数ヶ月程度休止期としてとどまります。その部位に新しい毛が生じると、休止期の毛は抜け落ちます。ヒトの毛では一本一本の毛周期が異なるため、毛の数が保たれています。毛周期に異常が起こると、脱毛の原因になります。もし毛周期をコントロールすることができれば、脱毛の治療にも役立つ事でしょう。今回ご紹介する論文は、毛周期をコントロールするメカニズムの一部を明らかにした論文です。
これまでの研究で、毛包幹細胞(※1)が、色素細胞、毛をとりまく神経や、毛の周りにある立毛筋と呼ばれる筋肉と密接に関与することが明らかにされてきました。しかし、毛包(※2)の周りにある脂肪細胞との関係は明らかではありませんでした。今回の論文では、皮膚に存在する脂肪前駆細胞が毛包幹細胞を維持し、毛が休止期から次の成長期へ進むのに重要な役割を果たすことが示されました。
まず筆者らは、毛包の再生に伴って、皮膚の脂肪層が厚くなることに着目しました。脂肪層が厚くなるのは、個々の脂肪細胞が大きくなるためではなく、脂肪前駆細胞が増加し、脂肪細胞の個数が増えるためである事がわかりました。さらに、この脂肪前駆細胞の増加は毛の休止期から成長期への移行と同時に起こるか、または少し先行する事も明らかにしました。
次に筆者らは、脂肪細胞に異常のある2種類の遺伝子変異マウスを用いて実験を行いました。脂肪生成の初期段階に異常があるために、脂肪前駆細胞が存在しないEfb1遺伝子欠損マウス(Ebf1-/-マウス)と、脂肪生成の最終段階が阻害されているために成熟脂肪細胞を欠いた変異マウス(Azip/F1マウス)を用いました。Azip/F1マウスでは、退行期の後期から休止期における毛のバルジ(※3)という部分に存在する幹細胞の活性化がみられるのに対し、Ebf1-/-マウスでは幹細胞の活性化がみられません。そこで、Azip/F1マウスの皮膚から脂肪前駆細胞を取り出し、Ebf1-/-マウスの皮膚へ移植しました。すると、Azip/F1マウスの脂肪前駆細胞が移植されたEbf1-/-マウスでは、毛の幹細胞が活性化し、休止期から成長期へと移行する像がみられたのです。すなわち、脂肪前駆細胞が毛の幹細胞を活性化し、休止期から成長期へ移行するのに重要な役割を果たす事が示されました。
では、脂肪前駆細胞が毛周期を制御するのに関与するシグナルは何でしょう?筆者らは、脂肪前駆細胞ではPDGF(※4)の発現が増加する事、成長期の毛乳頭にはPDGF受容体が活性化する事からPDGFに着目しました。筆者らは、Ebf1-/-マウスにPDGFでコートされたビーズを皮膚に注射すると、毛周期の異常が改善される事を示し、PDGFが毛周期を促進する因子として働く、と述べています。
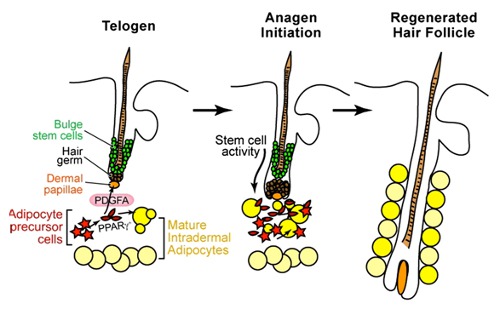
今回の結果から、毛の幹細胞を維持するのに脂肪前駆細胞が必要であり、そのシグナルにはPDGFが関与している事が明らかになりました。これまで知られていなかった、毛包と脂肪の関係が明らかにされた事はとても興味深いことです。発毛のメカニズムがまた一つ明かされたことになります。しかし、ヒトの発毛促進に応用するにはまだまだ検証が必要です。今回の結果はマウスの皮膚で示された結果なので、ヒトでも同様のメカニズムが働いているかはまだ不明です。また、成長期を開始させるシグナルはこれまでも報告されており、PDGF以外にもさまざまなシグナルが複雑に関係し合っている可能性が考えられます。今後も研究が進むことで、発毛の複雑なメカニズムが解明され、脱毛の治療につながっていくことが期待されます。
用語解説
- ※1 毛包幹細胞
毛を形作る角化細胞を供給する源の細胞。 - ※2 毛包
毛を囲むように入れている袋状の上皮性組織層。 - ※3 バルジ
毛包の隆起した部分.幹細胞が存在する部分でもある。 - ※4 PDGF(platelet-derived growth factor)
血小板由来成長因子。血液が凝固する時に血小板から放出される他、線維芽細胞、マクロファージ、平滑筋細胞、内皮細胞からも分泌される。PDGFは、組織が傷つくと放出されて、細胞遊走と細胞増殖を調節する事で,組織の修復、再生に関わる。
Copyright © Keio University. All rights reserved.